
Q.11. Describe the life cycle of Sphagnum. (2013)
Related Question-
Q. Write short note on Habit of Sphagnum.
Q. Describe the Gametophytic Phase of Sphagnum.
Q. Describe the Reproduction of Sphagnum.
Q. Describe the Sporophytic Phase of Sphagnum.
Q. Describe the structure of Mature Sporophyte.
Ans. Sphagnum: -
Habitat: - The invidual Sphagnum plants grow closely matted together forming extensive masses. The latter form a continuous and complete vegetative spongy cover over the surface of water in acid pools, ponds and lakes converting them to quaking bogs of peat lands. Sphagnum is also called peat moss.
Distribution: -
Sphagnum has a world-wide distribution. It occurs in all parts of the world except the Arctic region. It thrives in the tropics and extends through the temperate zone to the sub-arctic and the subantarctic region. Normally it grows on peat and in oligotrophic waters. In India it occurs in the Himalayas on the wet dripping rocks and on the sides of furrows cut by the fast flowing streams.
Habit of Sphagnum: -

Sphagnum plant generally is an erect perennial with various shades of green, pale green or yellow. Some species are of bright colours such as deep red (S. rubellum), orange brown, salmon to rose-pink (S. plumulosum). organge pink, etc. Sphagnum is counted among the largest mosses in point of size. As the plant grows its basal older parts die. The dead portions, accumulate from year to year as partially decomposed material which gradually fills the pond or the lake. These deposits may as well contain the remains of other plants growing there such as the sledges, heathers, cotton, grass etc. The water in which Sphagnum grows is often so acidic, that the decay of the accumulated plant tissue is very slow. These deposits of dead partially decomposed tissues are slowely compressed and hardened by the weight of fresh deposit above it. The compacted partially decomposed and hardened dead plant deposit is called the peat. The peat is a brown or dark-coloured spongy substance. It is rich in carbon. When air dried it is used as fuel.
Since Sphagunum is the chief component of peat it is often called the peat moss. It implies that it is not exclusively decomposed under the conditions which occur. in the surface, layers of peat bogs. Several explanations have been advanced for the apparent resistance to-decomposition shown by Sphagnum tissues. Czapek (1899) reported that the moss contains a phenolic compound termed sphagnol which inhibits microbial activity. Rumjantzewa (1939) suggested that Sphagnum cellulose is less easily broken down due to the presence of higher lignins. Lindberg and Theander (1052), and Farmer and Morrison (1964) confirmed the presence of unusual lignin containing hygroxyphenyl units in various species. Other workers have suggested that the physical environment is largely responsible for the tardy decomposition of dead Sphagnum tissues.
The life cycle of Sphagnum shows two distinct, alternating phases of generations, the gametophyte and the sporophyte phase.
Gamethophyte phase: -
It starts with the pore and consists of two stages, the juvenile stage and the leafy gametophyte stage.
Juvenile Stage: - It is also called the protonema and is formed by the germination of the spore. When young the protonema is a short filament of a few cells. By further growth and cell division the filament becomes a flat, green plate of cells. It is an irregulary lobed thallus-like structure on cell in thickness. It is fixed to the substratum by multicelluar rhizoids. The thallus-like protonema of Sphagnum moss recalls the thallose protonema of the leafy jungermanniales and thus points but the relationship of Sphagnum to a liverwort ancestor. From the margin of the lobed protonema arises the erect, leafy gametophyte called the Sphagnum plant. It is also called the gametophore. The stem of the young gametophore bears a few colourless rhizoids at its base. The rhizoids are delicate, multicellular, hair-like processes. The septa between the cells are oblique. As the gametophor grows the rhizoids soon disappear.The mature gametophytes of Sphagnum thus do not posses rhizoids.
(a) External Character: - The leafy gametophyte is a parenni1 plant. It has an up right stem. Individualiy the erect stem is weak and fragile. It gets support to grow upright from the neighbouring dense clups of stems. The stem often grows to a considerable length-a foot or more with a diameter up to 1.2 mm only. The mature gametophyte or adult plant lacks rhizoids but branches freely. The branching is lateral and the branches arise in fascicles. Both be stem and the branches are closed with small leaves. At the apex of the stem is a dense cluster of short, stout branches. These are of limited growth. This terminal cluster of closely set branches protects the apical bud and forms a conspicuous compact head of the plant. It is called the comma or commal tuft. As the apical bud grows these branches become displaced downwards.
Lower down on the stem are borne additional elongated branches. They usually occur in tufts or fasciles of 3 to 8 commonly five in the axil of every fourth leaf on the main stem.
(i) Divergent Branches: - These are usually short and stout branches which grow out laterally from the main axis. They extend outward in a horizontal position and sometimes slightly upward but usually become arched at the middle with their distal portions carving downward.
(ii) Flagelliform or Pendent Branches: - These are long, slender,descending branches which droop or hang down, often very close to and around the main stem. These pendent or decurrent branches act as water conductors. In the submerged species of Sphagnum (S. obesum) the drooping branches in cluster are rare. They are all of the divergent type.
Apical Growth: - The main stem and the branches grow by means of a three sided apical cells. It has three cutting faces. Each segment, cut off parallel to its flat face, divides by a peridinal wall into an outer cell and inner cell. The outer cell undergoes divisions to form the stem cortex and a single leaf. The inner cell gives rise to the central tissue of the stem. In this way each segment derived from the apical cell forms a leaf and the subtending portion of the stem. This explains the three arrangement in the younger portions of the stem. This arrangement is, however, soon replaced by more complex phyllotaxy.
Development of Leaf: - The leaf when young grows by means of an apical cell. It has two cutting faces. Segments are off alternately right and left parallel to its flat faces. This results in a young leaf consisting of a single layer of diamond shaped cells. They are all green and contain protoplasm and chloroplasts. Later the apical growth ceases. The further growth of the leaf is entirely basal. Each diamond-shaped green cell divides asymmetrically. It cuts off narrow daughter cells but on two sides only either right and tip side or leaf. The narrow daughter cells develop chloroplasts and remain green and alive. The larger mother cells lose chloroplasts and protoplasmic contents to become hyaline and empty. There empty cells develop entrance pores on their walls and spiral thickenings on the inner surface of the walls only.
Reproduction: -
Sphagnum reproduces by vegetative and sexual reproduction.
1. Vegetative reproduction: - Reproduction in sphagnum is principally. However, it lacks special structures for vegetative. The stem progressively disintegrate from the basal older parts upwards causing the branches to separate and develop into independent plants.
Innovation: - The common vegetative method is by the formation of innovation. Occasionally one of the branches in the axillary cluster may turn upward. It grous more vigorously than the others and continues its upward growth. This long, upright branch takes on all the characteristics of the main axis. It is called an innovation. Sooner or later the innovations become separated from the main stem by progressive dying of their basal, older parts. The detached innovation establishes itself as an independent individual. Vegetative propagation by this method is a very effective means of multiplication in Sphagnum. It is mainly responsible for its occurrence in extensive masses.
Regeneration: - Sphagnum like other bryophytes has great power of regeneration. It is attributed to the physiological adaptations which allow suspension of metabolism during periods of desiccation and its resumption when water is once more available.

Gemmae: - These structures were previously unknown for sphagnales. Redhead (1981) discovered gemmae in S. capillaceum. The gemmae are subglobose and double-walled. Usually they occur in pairs, sometimes in clusters or even singly. On germination the gemma produces a small, uniseriate
filamentous protonema prior to prothallus development.
2. Sexual Reproduction: - The plant may be monoecious or diocecous but the sex organs always occur on separate and independent branches. They are formed in autumn on special, short, densely leafy and slightly modified branches. The sexual branches either occur in the terminal branch cluster, the comma or lower down on the stem. In some species the antheridial and archegonial branches are borne on the same plant in others on separate plants. Even in the monoecious species the two kinds of sex organs never occur on the same branch. The antheridial branches appear first. The sex organs are formed in abundance. Paraphyses are always absent. Benson-Evans (1964) reported that S. plumulosum is perhaps the only moss studied in which initiation of sexuality is induced by short days. In all others temperature seems to be the critical factor.

(a) Antheridial Branches: - The antheridial barnches are usually shorter but stouter than the vegetative branches. They are spindle-shaped and like small catkins. They are strongly pigmented and often densely clothed with red, purple, brown or yellow leaves generally smaller than the falioge leaves.

(i) Development and Structure: - The antheridia occur singly in the axils of leaves towards the tip on antheridial branches. The mature autheridium consists of a long stalk and globose body. The stalk may be as long as the body of the antheridium and consists of 2-4 vertical rows of cells. The body of the antheridium is bounded by a jacket layer of sterile cells. It is known as the antheridial wall. Hofmeister (1862) reported the presence of a glossy transparent tough cuticle layer on the living autheridium of Sphagnum. Manton (1954) confirmed it with the help of electron microscope. He reported that the cuticle forms a very conspicuous layer on the outer side. The one cell thick antheridial wall encloses a mass of androcyte mother cells. Each androcyte mother cell divides into two androcyte. The division is not diagonal. The androcytes metamorphose into sperms. Each sperm is an elongated, spirally coiled structure furnished with two flagella. The number of coils is 2 to 3. The flagella are inserted at the end which is cytoplasmic in origin and called the flagellophore. The greater part of the body of the sperm is nuclear in origin. To its posterior end is attached a vesicle-like structure consisting of a cytoplasmic matrix.
(ii) Dehiscence: - The mature antheridium wall absorbs the water and dehiscence by splitting of wall into several irregular lobes. The jacket cells of the swollen antheridium undergo irregular separation at its distal end. The antheridium thus opens at its apex by a number of irregular lobes. The ejection of sperms is further faciliated by swelling of the jacket cells which cause the lobes to bend outwards away from the aperture. The formerly convex outer surfaces of these cells become concave, and the inner surfaces previously concave due to internal pressure of spermatocytes become convex. With the, dissolution, of the androcyte vesicles, the sperms are liberated. They swim about actively and reach the archegonium.

(b) Arehegonial branches: - The female branches are very short and more or less globular, bud-like aggregates of bracts and archegonia. In many species the archegonial branch occurs just below the apex of the stem within the terminal branch cluster. It bears, leaves larger than the normal. These leaves are rich in chloroplasts and have fewer fibres in the hyaline cells. The archegonial branches are thus more deeply pigmented than the vegetative branches in the cluster. The leaves increase in size towards the apex where a number of them form the perichaetium. The perichaetium surrounded and protects the archegonia.
(1) Development and Structure: - The archegonia are terminal on the specialized archegonal branches. They usually occur in small groups. Typically there are three archegonia in the group. The number, however, varies from 2 to 5. The central or the middle arhegonium in the cluster grows directly from the apical cell of the branch. It is called the primary archegonium and is the firm to be formed. The others in the cluster are called the secondary archegonia. They are developed from the last segments cut off by the apical cell. The secondary are thus formed about the base of the primary archegnium.
The mature archegonium is relatively a large structure. It is stalked. The stalk is fairly long. The. body of the archegonium consists of a long twisted neck and a massive venter. The neck consists of six vertical rows of neck cells. The neck canal contains numerous (usually 8 or 9) neck canal cells. The, venter and the lower portion of the neck is 2 to 3 layers thick of cells. The venter cavity contains a small ovoid egg. it is about the same size as the ventral canal cell which lies above it.

Fertilization: - In S.subsecundyn the ventral canal cell persists and fuses with the egg and forms the zygote. Fertilisation, however, occurs in the same manner as in the other bryophytes. The axial row of cells in the archegonium, except the egg, disintegrates to form a passage way for the sperms. The released sperms swim to the archegonium. Some of them find their way into the neck cannal and swim down to the egg. One of them fuses with the egg to for the diploid zygote.
The diploid zygote is thus the pioneer structure of this generation. The zygote encased in the venter, increase in size, secretes a wall around it and starts dividing. By repeated division it develops into an embryo which by further division, differentiation and growth forms the young sporogonium or sporophyte. Usually the zygote on one archegonium in the cluster develops into the sporogonium. The others persist for sometime and finally abort. One wet moors and peat bogs fruiting species of Sphagnum in June can be seen bearing pale greenish young sporogonia (sporophytes) amongst the leaves of the terminal clusters of branches. By the middle of July they mature and each is seen elevated on a short, cylindrical and leafless stalk, the pseudopodium which is of gametophyte origin. The mature sporogonia are brown or dark brown in colour and ready for spore dispersal.

The ripe sporogonium or sporophyte consists of a foot and a capsule. The two are linked by a small, narrow, neck-like constriction which represents the suppressed seta.
Mechanism of dehiscence and dispersal of spores: -
The dehiscence of the capsule is by an explosive mechanism. The dispersal occurs in a sunny day. The capsule wall becomes dry and shrinked. It causes the spherical capsule to become cylindrical and thus compressing.
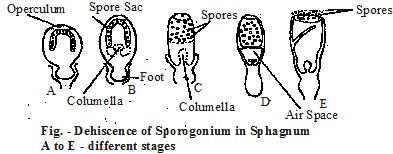
the enclosed air of spore-sac. The pressure flicks away the operculum with audible sound and accounts for the rupture of spore sac. It is called “air gun” mechanism of spore discharge. in submerged plants, a hole is formed at the base of capsule during spore dispersal.
Spore Structure: - The released spores are the pioneer structures of the future young gametophyte. They are tetrahedral with a prominent tri-radiate mark and yellow to brown in colour. Each spore is a uninucleated tiny mass of protoplast protected by the surrounding spore wall. Spore cytoplasm contains one to several large oil droplets (S. palustre). The spore has two layered wall, outer exine (exosporium) and inner intine, (endosporium). The exine is brown smooth granular or papillate and itine is thin.
Germination of Spores: - The spores remain viable for six months or so under dry conditions. Under favourable conditions the spore germinates within a weak’s time. It absorbs water and swells up slightly (A). The exposporium ruptures. The endosporium grows through the split as a short protuberance, The germ tube which turns green. The green, tube grows and divides by transverse wall and form 2-4 celled filament or protonema. At this stage the terminal cell of the filament begins to function as an apical cell. It has two cutting faces. Segments are cut off parallel to these alternately right and left. Eventually a flat green protonema is formed.

The Protonema: - It consists of photosynthetic cells. They are arranged in the form of a plate, cell thick. During further growth the apical cell becomes inactive and indistinguishable. Certain marginal cells of the plate-like protonema undergo anticlinal and periclinal divisions. As a result the primary protonema becomes an irregularly lobed, green plate, one cell in thickness. It is fixed to the substratum by colourless, multicellular rhizoids with oblique septa. The rhizoids are unbranched. The ruptured exspore persists at the marginal cells near the base of the primary protonema arises the erect, leafy gametophore. The cell by growth and segmentation form a tiny bud. Soon a tetrahedral apical cell becomes stablisbed in the bud. It cuts off segments like the apical cell of the adult gametphore. As a result an upright, apically growing shoot of Sphagnum is formed.
